
Растения в процессе своего роста и развития часто испытывают воздействие неблагоприятных факторов внешней среды, к которым относят температурные колебания, засуху, избыточное увлажнение, засоленность почвы и т.д. Защита от неблагоприятных факторов среды у растений обеспечивается структурными приспособлениями (мелколиственность или отсутствие листьев, густое опушение, погруженность устьиц, наличие сочных листьев и стеблей, сохраняющих резервы воды и др.), особенностями анатомического строения (кутикула, корка, механические ткани и т.д.), специальными органами защиты (жгучие волоски, колючки), двигательными и физиологическими реакциями, выработкой защитных веществ (смол, фитонцидов, токсинов, защитных белков) (Володько, 1983).
Первым барьером в адаптационной системе растений к внешней среде является эпидерма. Особенно значимый структурный показатель листа – устьичный аппарат. Из всех эпидермальных производных количественные характеристики устъичного аппарата наиболее фенотипически пластичны и чувствительны к изменению климатических условий (Спесивцева, Александрович, 1989). Следует отметить малочисленность работ, посвященных изучению эпидермальных структур, как показателя адаптивности с экологической точки зрения. В трудах дальневосточных ученых (Ворошилова, 1980; Чернышев, 1993, 1996, 2001) описаны изменения эпидермальных производных для некоторых хвойных растений местной флоры.
С учетом малой изученности комплексного воздействия среды на эпидерму листа растений значительную актуальность представляет получение информации по морфологии устьичного аппарата культивируемых в нашей стране комнатных растений, как представителей флоры разных природных зон (Чернышев, 1996).
Цель настоящей работы – провести сравнительный анализ особенностей морфологии устьичного аппарата комнатных растений.
В связи с поставленной целью сформулированы следующие задачи:
1) определить климатические особенности природных ареалов комнатных растений;
2) исследовать морфологические особенности строения устьичного аппарата у выбранных комнатных растений;
3) провести сравнительный анализ морфологии устьичного аппарата комнатных растений.
Обзор литературы
Листья и молодые зеленые побеги, как чехлом, покрыты однослойной первичной покровной тканью – эпидермой. Изредка эпидерма многослойна. Такая многослойная эпидерма обнаружена в листьях известного комнатного растения фикуса (Ficus elastica). Эпидерма – сложная ткань, поскольку ее клетки различаются по форме и отчасти по функциям. В частности, резко отличаются клетки, образующие устьица, и клетки трихом.
Наружная поверхность клеток эпидермы часто покрыта слоем кутикулы или, реже, восковым налетом различной толщины. Кутикула может достигать значительной толщины, особенно у растений засушливых мест обитания. Нередко ее поверхность покрыта различного рода складками или бородавчатыми выростами. Исключая устьичные щели, клетки эпидермы плотно сомкнуты, т.е. отсутствуют межклетники. Главная функция эпидермы – регуляция газообмена и транспирации, т.е. испарения воды растением. Газообмен и транспирация осуществляются преимущественно через устьица, но частично и через кутикулу. Кроме того, через поры и тяжи пектиновых веществ в наружных стенках клеточных оболочек эпидермы проникают вода и неорганические питательные вещества, что особенно характерно для водных растений.
Иногда эпидерма выполняет необычные для этой ткани функции – такие, как фотосинтез (у части водных растений), запасание воды (у некоторых пустынных растений) или секрецию веществ вторичного метаболизма (ряд эфирномасличных) (Бавтуто, 2001).
Характер клеток эпидермы различен, большинство, получившее название основных клеток эпидермы, отличается разнообразием очертаний. Боковые стенки, как правило, извилисты, что повышает плотность их сцепления друг с другом, реже прямые. Эпидермальные клетки осевых органов и листьев многих однодольных сильно вытянуты вдоль оси органа.
Через устьица проходит чрезвычайно интенсивная диффузия водяного пара, кислорода и углекислого газа (СO2).
Каждое устьице состоит из пары замыкающих клеток и устьичной щели, которая представляет собой межклетник. Замыкающие клетки отличаются от окружающих их обычных эпидермальных клеток своей формой и наличием хлоропластов. На поперечном срезе, сделанном через замыкающие клетки, ясно видно, что устьичная щель ведет в более или менее хорошо выраженное межклеточное пространство, называемое подустьичной или дыхательной полостью. Она сообщается с другими межклетниками, пронизывающими ткань листа, что облегчает их газообмен с окружающей воздушной средой.
При недостатке воды и света, а также при низких (или, наоборот, очень высоких) температурах замыкающие клетки, смыкаясь, закрывают устьичную щель и транспирация частично или полностью прекращается. Но в тех случаях, когда нет недостатка в воде и активные процессы фотосинтеза вызывают необходимость газообмена со средой, устьица открываются. Раскрывание и закрывание устьиц представляет собой чрезвычайно важное в жизни высших растений явление, которое еще далеко не полностью изучено. Но со времен немецкого ботаника С. Швенденера (1829–1919) мы знаем, что основным фактором является здесь изменение тургора, вызываемое соответственно повышением или понижением осмотического давления внутри замыкающих клеток.
Наличие устьиц является одним из наиболее характерных признаков высших растений, начиная с древнейших известных их представителей – риниофитов. Устьиц, как правило, нет в покровных тканях корней. Как результат редукции, устьица отсутствуют у многих погруженных в воду растений, многих санрофитов и паразитов, а также у большинства мхов и печеночников. В последнем случае устьица редуцировались в результате потери спорофитом способности к самостоятельному существованию. Фотосинтетическая его активность поэтому стала минимальной. Но наличие рудиментарных устьиц у спорофитов ряда моховидных говорит в пользу предположения, что у их предков устьица были нормально развиты и играли важную физиологическую роль.
Между бесцветными клетками этой защитной ткани расположены устьица, образованные двумя полулунными клетками, между которыми остается межклеточное пространство – устьичная щель. Изогнутость этих клеток изменяется в зависимости от условий окружающей среды: в первую очередь от обеспечения растений водой, отчего образуемое ими отверстие может расшириться, сузиться или почти совсем закрыться. Обычно большая часть устьиц находится на нижней стороне листа. Клетки, образующие устьица, содержат хлоропласты, участвующие в процессе фотосинтеза.
Через устьица внутрь листа свободно проникает углекислый газ, кроме того они регулируют испарение воды растением. Эти отверстия имеют микроскопическую величину (около 0,00005 мм2), но на каждом листе их очень много.
Число и распределение устьиц, типы устьичных аппаратов широко варьирует у различных растений. У различных растений насчитывают на 1 мм 2 поверхности листа от 40 до 700 и более устьиц. Средний по величине лист капусты или подсолнечника имеет свыше 10 млн устьиц. Несмотря на такое большое количество устьиц, общая площадь их отверстий составляет не более одной сотой доли всей поверхности листа. Однако малая величина устьиц не является препятствием для проникновения углекислоты внутрь листа, так как все газы, в том числе и углекислый, обладают способностью проникать через малые отверстия с большой скоростью.
Сравнительное изучение строения и развития устьиц имеет большое значение для систематики и палеоботанических исследований.
Типы устьиц. Разнообразие типов устьичного аппарата привело к необходимости их классификации. Разные типы устьичного аппарата различаются прежде всего наличием побочных клеток, их числом и расположением по отношению друг к другу и к замыкающим клеткам (рис. 1). Наряду с классификациями морфологических или топографических типов устьичного аппарата за последнее десятилетие появились классификации онтогенетических их типов. Первая попытка классифицировать онтогенетические типы была предпринята индийским ботаником Д. Пантом в 1965 г. Он выделил три крупные категории устьичных аппаратов в зависимости от того, происходят ли клетки, окружающие устьице, из той же самой материнской клетки, что и устьице, или из других клеток:
1) мезогенный (от греч. mesos – средний и genos – происхождение) устьичный аппарат, у которого все побочные клетки или одна кольцеобразная побочная клетка происходит из той же самой материнской клетки, что и замыкающие клетки;
2) мезоперигенный устьичный аппарат, у которого только одна из окружающих клеток образуется таким путем, остальные образуются из других клеток;
3) перигенный (от греч. peri – около, кругом) устьичный аппарат, у которого ни одна из окружающих клеток не происходит из той же самой материнской клетки, что и замыкающие клетки.
Таким образом, в основу классификации Панта был положен способ развития устьичного аппарата без учета типов взрослых структур, тогда как классификация морфологических типов строилась исключительно на расположении побочных клеток в зрелом состоянии, при полном игнорировании способа развития устьичного аппарата. Только появление в 1973 г. классификации онтогенетических типов устьичного аппарата, разработанной Е. Фринс-Классенсом и В. Ван Коттемом, устранило разрыв между морфологической и онтогенетической классификациями (Тахтаджян, 1982).
Методы и материалы исследования
Для определения естественного природного ареала и климатических условий произрастания были использованы следующие литературные источники – Бавтуто Г.А., Еремин В.М., Жигар М.П. Атлас по анатомии растений. 2001, Володько И.К. Микроэлементы и устойчивость растений к неблагоприятным условиям. 1983 г., Ворошилова Г.И. Характеристика эпидермы листа дальневосточных хвойных / Г.И. Ворошилова II Ботанические исследования на Дальнем Востоке. 1980, Головкин Б.Н., Чеканов В.Н., Шахова Г.И. Комнатные растения. 1989, Сааков С.Г. Оранжерейные и комнатные растения. 1983, Тахтаджян А.Л. Жизнь растений. В 6-ти томах. Том 6, Цветковые растения. 1982, http://dic.academic.ru/,Чернышев В.Д. Очерк экофизиологии лесных растений Дальнего Востока / В.Д. Чернышев II Биологические исследования в естественных и культурных экосистемах Приморского края. Сборник научных трудов. 1993, Чернышев В.Д. Принципы адаптации живых организмов (экологический аспект) / В.Д. Чернышев. 1996, Чернышев В.Д. Об особенностях строения и развития сложных лесов Дальнего Востока. 2001.
Микроскопическое исследование проводилось лабораторным методом с использованием цифровой фотографии на микроскопе «JJ – Optics Digital Lab» на увеличениях до 400Х в проходящем свете.
Тип устичного аппарата определяли согласно классификации по морфологическим особенностям устьичного аппарата (Приложение I рис. 1, табл. 1)
Таблица 1
Классификация по морфологическим особенностям устьичного аппарата описанная А.Л. Тахтаджян (Тахтаджян А.Л., 1982)
|
№ п/п |
Типы устьичного аппарата |
Морфологические особенности устьичного аппарата |
|
1 |
Аномоцитный (от греч. anomos – беспорядочный) |
Замыкающие клетки устьиц окружены клетками, не отличающимися от остальных клеток эпидермы. Во всех группах высших растений за исключением хвощей |
|
2 |
Перицитный (от греч. peri – около, вокруг) |
Замыкающие клетки полностью окружены одной побочной клеткой. Только у папоротников |
|
3 |
Полоцитный (от греч. polos – полюс) |
Замыкающие клетки окружены одной побочной клеткой не полностью: к одному из устьич-ных полюсов примыкают одна пли две эпидермальные клетки. Главным образом у папоротников |
|
4 |
Диацитный (от греч. Dia – врозь, через) |
Замыкающие клетки окружены парой побочных клеток, общая стенка которых находится под прямым углом к замыкающим клеткам. У папоротников и цветковых |
|
5 |
Парацитный (от греч. para – рядом) |
каждая из замыкающих клеток устьиц сопровождается одной или более побочными клетками, расположенными параллельно замыкающим клеткам. У папоротников, хвощей, цветковых и гнетопсид |
|
6 |
Анизоцитный (от греч. anisos-неравный) |
Замыкающие клетки устьиц окружены тремя побочными клетками, из которых одна заметно крупнее или меньше двух других. Только у цветковых |
|
7 |
Тетрацитный (от греч. tetra – четыре) |
Замыкающие клетки окружены четырьмя побочными клетками, из которых две латеральные и две полярные. У цветковых, главным образом у однодольных |
|
8 |
Ставроцитный (от греч. stauros – крест) |
Замыкающие клетки устьиц окружены четырьмя (иногда тремя или пятью) одинаковыми, более или менее радиально вытянутыми побочными клетками, антиклинальные стенки которых расположены накрест но отношению к замыкающим клеткам. У папоротников, изредка у цветковых |
|
Окончание табл. 1 |
||
|
№ п/п |
Типы устьичного аппарата |
Морфологические особенности устьичного аппарата |
|
9 |
Энциклоцитный (от греч. kyklos – колесо, круг) |
четыре (иногда три) или более побочных клеток образуют узкое кольцо вокруг замыкающих клеток. У папоротников, голосеменных и цветковых |
|
10 |
Актиноцитный (от греч. aktis – луч) |
пять или более радиально вытянутых побочных клеток располагаются вокруг замыкающих клеток. Только у цветковых |
Глава 3. Результаты и обсуждение
Нами были исследованы устьичные аппараты 25 видов комнатных растений (Приложение I) и определены их типы (Таблица 2). Большинство наших комнатных растений – выходцы из тропических и субтропических зон Азии, Африки, Европы, Америки и Австралии (Приложение II) (Ньюмен,1989).
На основании результатов микроскопического исследования эпидермы растений можно выделить ряд анатомических, признаков общих для всех исследуемых видов: наличие замыкающих клеток, устьичной щели, клетки эпидермы плотно сомкнуты, т.е. отсутствуют межклетники, у основных клеток эпидермы, боковые стенки, как правило, извилисты, реже прямые.
Таблица 2
Родина произрастания растений и типы устьичного аппарата
|
№ п/п |
Название растения |
Родина, природная зона, условия произрастания |
Тип устьичного аппарата |
|
1 |
Антуриум андре |
Колумбия, Эквадор, влажные тропические леса, 1000 м. над уровнем моря |
Анизоцитный |
|
2 |
Аспарагус Шпренгера |
Восточная и Южная Африка провинция Наталь в ЮАР, субтропические и тропические леса, влажные горные склоны |
Тетрацитный |
|
3 |
Бегония вечноцветущая |
Бразилия, Южная и Восточная Азии, |
Диацитный |
|
4 |
Гемантус белоцветковый (тещин язык) |
Южная Африка, на острове Сакотра, тропики, скалистые склоны гор, заросли кустарников |
Тетрацитный |
|
5 |
Гибискус (розан китайский) |
Тропики, Африка, остров Мадагаскар и Южно-восточная Азия, родина – Юго-Восточная Азия и Полинезия, сухие субтропики |
Анизоцитный |
|
6 |
Диффенбахия пятнистая |
Родина – Центральная и Южная Америка, |
Парацитный |
|
7 |
Зигокактус усеченный |
Восточная Бразилия, влажные тропические леса |
Парацитный |
|
8 |
Каланхое Дегремона |
Ю. Америка, Ю. Африка, Мадагаскар, тропики |
Диацитный |
|
9 |
Кливия киноварная |
Юго-Восточная Африка провинция Наталь в ЮАР, влажные тропические леса от прибрежья поднимаясь в горы до высоты 600–800 м над уровнем моря |
Тетрацитный |
|
10 |
Кодиеум пестрый (Кротон) |
Индия, острова Малайского архипелага, |
Парацитный |
|
11 |
Молочай блестящий (эуфорбия) |
Остров Мадагаскар, сухие субтропики |
Парацитный |
|
12 |
Монстера неравнобокая |
От Мексики до Бразилии, родина – Бразилия, влажный тропический лес |
Парацитный |
|
13 |
Пеларгония зональная |
Южная Африка, сухие субтропики |
Ставроцитный |
|
14 |
Пеперомия клузиелистная |
Антильские острова, Венесуэла, |
Перицитный |
|
15 |
Сансевиерия трехполосчатая |
Западная Африка, бассейн реки Конго, |
Перицитный |
|
16 |
Узумбарская фиалка |
В. Африка, горные районы, сухие субтропики |
Перицитный |
|
Окончание табл. 2 |
|||
|
№ п/п |
Название растения |
Родина, природная зона, условия произрастания |
Тип устьичного аппарата |
|
17 |
Сеткреазия пурпурная |
Мексика, высокогорный районы, сухие тропики |
Парацитный |
|
18 |
Сингониум ножколистный |
Центральная и Южная Америка |
Ставроцитный |
|
19 |
Спатифиллюм Уоллиса |
Колумбия, влажные тропические леса, |
Аномоцитный |
|
20 |
Толстянка древовидная |
Южная Африка, Мадагаскаре, |
Перицитный |
|
21 |
Фикус каучуконосный |
Северо-Восточная Индия, Непал, Бирма, |
Перицитный |
|
22 |
Филодендрон лазящий |
Центральная Америка и низменные пространства Южной Америки, влажный тропический лес |
Парацитный |
|
23 |
Хлорофитум хохлатый |
Южная Африка, провинция Наталь в ЮАР, влажные субтропики |
Аномоцитный |
|
24 |
Хойя мясистая |
Индия, Южный Китай, |
Энциклоцитный |
|
25 |
Шеффлера восьмилисточковая |
Восточная Азия до Китая, |
Парацитный |
|
|
|
|
|
|
Диффенбахия пятнистая |
Зигокактус усеченный |
Кодиеум пестрый |
Молочай блестящий |



|
|
|
|
|
|
Монстера неравнобокая |
Сеткреазия пурпурная |
Филодендрон лазящий |
Шеффлера восьмилисточковая |


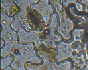

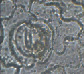
Анализ микрофотографий и родины произрастания растений указывает на то, что строение устьичного аппарата растения не будет зависеть от климатических условий произрастания растения (Приложение II). Так один исследуемый тип устьичного аппарата встречается не в одной природной зоне. Например парацитный тип, где каждая из замыкающих клеток устьиц сопровождается одной или более побочными клетками, расположенными параллельно замыкающим клеткам, имеют некоторые растения тропической зоны, такие как – Кодиеум пестрый (Кротон), Сеткреазия пурпурная, Шеффлера восьмилисточковая; влажной тропической зоны – Диффенбахия пятнистая, Зигокактус усеченный, Монстера неравнобокая; сухих тропиков – Диффенбахия пятнистая, Монстера неравнобокая, Филодендрон лазящий; субтропиков – Молочай блестящий (эуфорбия) (Таблица 1, схема 5).
Или к примеру – тетрацитный тип, в котором замыкающие клетки окружены четырьмя побочными клетками, из которых две латеральные и две полярные, имеют растения тропической зоны – Гемантус белоцветковый (тещин язык),субтропической зоны – Аспарагус Шпренгера, Кливия киноварная (Таблица 1, схема 7).



Гемантус белоцветковый Аспарагус Шпреигера Кливия киноварная
БЕГОНИЯ (Begonia), род растений семейства бегониевых. Многолетние травы, реже полукустарники, кустарники, лианы и эпифиты. Около 1000 видов, выходцы из тропик и субтропик (кроме Австралии).
Таблица 3
Типы устьичного аппарата у растений рода Бегония
|
№ п/п |
Вид растения |
Тип устьичного аппарата |
|
1 |
Бегония Black major |
Анизоцитный |
|
2 |
Бегония Точечная |
Парацитный |
|
3 |
Бегония Борщевиколистная |
Ставроцитный |
|
4 |
Бегония Бауэра |
Устья расположены группами Анизоцитный |
|
5 |
Бегония Воротничковая «Крипса» |
Ставроцитный |
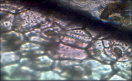
У растений относящихся к одному роду Бегония типы устьичного аппарата разные. Таким образом тип устьичного аппарата не является родовым признаком.
|
|
|
|
|
Бегония Black major Анизоцитный |
Бегония Точечная Парацитный |
БегонияБорщевиколистная Ставроцитный |


|
|
|
|
Бегония Бауэра Бегония Устья расположены группами Анизоцитный |
Воротничковая «Крипса» Ставроцитный |


Выводы
1. Большинство наших комнатных растений – выходцы из тропических и субтропических зон Азии, Африки, Европы, Америки и Австралии.
2. У исследуемых комнатных растений определено 8 типов устьичного аппарата.
3. Самый распространенный тип устьичного аппарата – парацитный.
Таким образом, наша гипотеза не подтвердилась. Тип устьичного аппарата не зависит от условий обитания и не является родовым признаком.
Приложение I
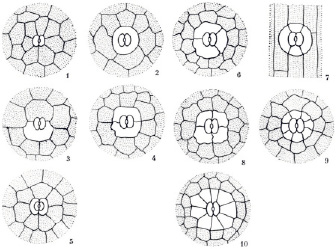
Рис. 1. Типы устьичного аппарата описанные А.Л. Тахтаджян ( Тахтаджян А.Л., 1982)
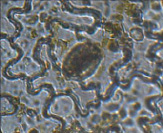
Антуриум андре – анизоцитный

Аспарагус Шпреигера – тетрацитный
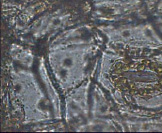
Бегония вечноцветущая – диацитный
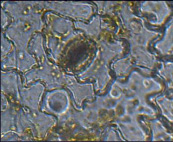
Гемантус белоцветковый
(тещин язык) – тетрацитный
Гибискус (розан китайский) – анизоцитный
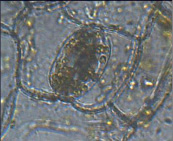
Диффенбахия пятнистая – парацитный
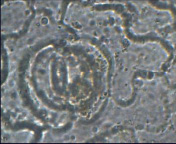
Зигокактус усеченный – парацитный
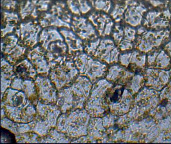
Каланхое Дегремона – диацитный

Кливия киноварная – тетрацитный

Кодиеум тертый кротон – парацитный
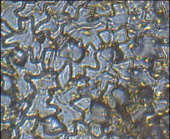
Молочай блестящий эуфорбия – парацитный
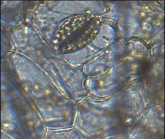
Монстера неравнобокая – парацитный
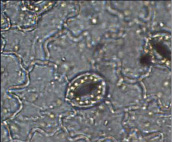
Пеларгония зональная – ставроцитный
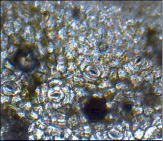
Пеперомия клузнелистная – перицитный

Сансевиерия трехполосчатая – перицитный

Сеткреазия пурпурная – парацитный
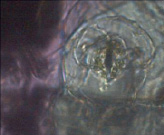
Узумбарская фиалка (Сенполия) – перицитный

Сингониум ножколистный – ставроцитный
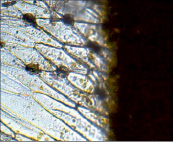
Спатифиллюм Уоллиса – аномоцитный

Толстянка портулаковая – перицитный
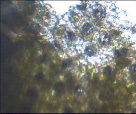
Фикус каучуконосный – перицитный
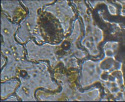
Филодендрон лазящий – парацитный

Хлорофитум хохлатый – аномоцитный
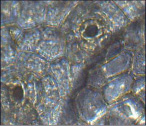
Хойя мясистая – энциклоцитный

Шеффлера восьмилисточковая – парацитный
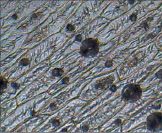
Рис. 2. Микрофотографии устьичных аппаратов исследуемых комнатных растений (окончание рисунка)
Приложение II
Биогеографическая классификация и типы устьичных аппаратов
исследуемых комнатных растений
|
Природная зона |
Название растений |
Типы устьичных аппаратов |
|
Тропики |
Гемантус белоцветковый (тещин язык), Каланхое Дегремона, Кодиеум пестрый (Кротон), Пеперомия клузиелистная, Сеткреазия пурпурная, Спатифиллюм Уоллиса, Фикус каучуконосный, Шеффлера восьмилисточковая |
парацитный тетрацитный диацитный перицитный аномоцитный |
|
Влажные |
Антуриум андре, Диффенбахия пятнистая, Бегония Хойя мясистая |
парацитный анизоцитный диацитный ставроцитный |
|
Сухие тропики |
Диффенбахия пятнистая, Монстера неравнобокая, Филодендрон лазящий |
Парацитный |
|
Субтропики |
Аспарагус Шпренгера, Гибискус (розан китайский), Кливия киноварная, Молочай блестящий (эуфорбия), Пеларгония зональная, Узумбарская фиалка (Сенполия), Толстянка древовидная, Хлорофитум хохлатый |
парацитный анизоцитный тетрацитный ставроцитный перицитный аномоцитный |
Библиографическая ссылка
Артюшин К.Е. СРАВНИТЕЛЬНЫЙ АНАЛИЗ МОРФОЛОГИИ УСТЬИЧНОГО АППАРАТА КОМНАТНЫХ РАСТЕНИЙ // Старт в науке. 2019. № 2-3. ;URL: https://science-start.ru/ru/article/view?id=1482 (дата обращения: 20.06.2026).